Fyziológia drevín: fotosyntéza, dýchanie a vodný režim
Ako stromy vyrábajú energiu, dýchajú a hospodária s vodou? Podrobný sprievodca fyziológiou drevín od fotosyntézy cez Krebsov cyklus až po vodný potenciál buniek.
Čo je fotosyntéza a prečo je základom života na Zemi
Základy fyziológie drevín
Fotosyntéza

Fotosyntéza (z gréc. phos, photós = svetlo) je anabolický, skladný dej, prebiehajúci u autotrofných, fotosyntetických organizmov (fotosyntetických baktérií a rias a najmä zelených rastlín), na ktorých činnosti závisí existencia ďalších dejov v živých organizmoch, t. j. respirácia a syntézy všetkých typov organických látok biosféry našej planéty. Fotosyntéza umožňuje zachytávať a viazať elektromagnetickú energiu slnečného žiarenia do energie chemických väzieb, využiť ju na rozbitie molekuly vody (fotolýza), a tým obohacovať atmosféru o kyslík, a napokon umožňuje zabudovať atómy uhlíka, kyslíka (zo vzdušného CO₂) a vodíka (z vody, H₂O) do molekúl jednoduchých cukrov. Každému mólu prijatého oxidu uhličitého zodpovedá zisk 477 kJ potenciálnej energie.
Následnými procesmi vytvárajú rastliny malé molekuly aminokyselín, nukleotidov a mastných kyselín, ktoré sú v rôznych metabolických dráhach začleňované do bielkovín (proteínov), nukleových kyselín, polysacharidov a lipidov, t. j. základov nadmolekulárnych funkčných a štrukturálnych systémov. Tieto zložité syntetické procesy však vychádzajú až z respirácie, procesu charakteristického pre všetky živé organizmy, resp. ich živé bunky. Organické látky, primárne pochádzajúce z fotosyntézy, slúžia ako molekuly potravy pre heterotrofy, neschopné žiť len z anorganických zdrojov (herbivory, omnivory, parazity, saprofyty). Z tohto uhla pohľadu sú fotosyntetizujúce organizmy producentmi organickej hmoty a na nich závisia všetky ostatné formy života, t. j. konzumenti (živiaci sa rastlinnou aj živočíšnou potravou) a reducenti (živiaci sa rozkladom, mineralizáciou odumretých tiel všetkých skupín organizmov).
Fotosyntéza zahŕňa fotochemické procesy, ktoré prebiehajú za prítomnosti svetla (tzv. primárne procesy), enzymatické procesy, ktoré svetlo nevyžadujú (t. j. temnnostné, sekundárne reakcie), a procesy difúzie, ktoré umožňujú výmenu oxidu uhličitého a kyslíka.
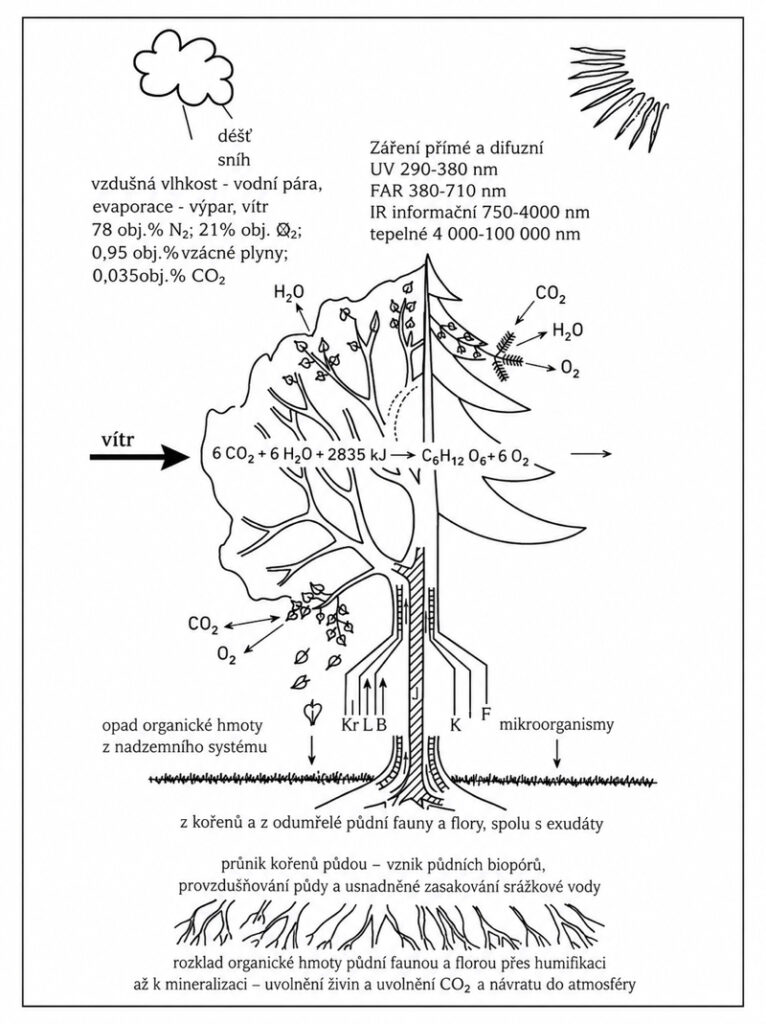
Schéma hlavných fyziologických funkcií nadzemného a podzemného systému. Voda a minerálne živiny sú koreňmi získavané z pôdy a živou časťou dreva (bielom – B) transportované do nadzemného systému. V listoch sú voda, minerálne živiny a najmä CO₂, prijatý z atmosféry, spracované v procese fotosyntézy (za vstupu žiarivej energie – FAR). Ohrevom listov a vetrom sa urýchľuje výpar vody – transpirácia, ale zároveň je k listom privádzaný CO₂ a O₂. Fotosyntáty z listov sú rozvádzané lýkom (L) do miest spotreby – rastových vrcholov, kambia (K), felogénu (F) a najmä do koreňového systému. Všetky časti stromu, resp. živé bunky, dýchajú, tj. spotrebúvajú fotosyntáty a uvoľňujú CO₂. Vnútorná časť kmeňa, vetiev, koreňov je tvorená odumretým drevom (jadrom – J) bez fyziologických aktivít. Počas rastu stromov dochádza aj k odumieraniu ich jednotlivých častí – lístia, koreňov, vetiev a krycích pletív vrátane kôry (Kr). Týmto opadom sa pôda obohacuje o organicky viazaný uhlík. Umožňuje život pôdnej mikroflóry a fauny vrátane mykorhitických húb, podieľajúcich sa na humifikácii až mineralizácii, tj. uvoľnení živín a uvoľnení CO₂ .

Fotosyntetické pigmenty: chlorofyly a karotenoidy v listoch drevín
Na zachytenie slnečnej energie slúžia fotosynteticky aktívne pigmenty, ktoré sa u drevín vyskytujú v bunkách listového mezofylu, v primárnej kôre listových stopiek a stoniek, resp. vetiev, a v ostatných zelených častiach nadzemného systému:
1. chlorofyly: vyššie rastliny obsahujú chlorofyl a a chlorofyl b. Chlorofyly sú porfyríny, deriváty tetrapyrolu, spojené s komplexne viazaným katiónom Mg²⁺. Sú veľmi podobné ľudskému hemoglobínu červených krviniek, ktorý však obsahuje atóm železa. Ich farbu a funkciu podmieňuje stavba molekúl vrátane ich usporiadania v bielkovinnom komplexe chloroplastových membrán (tylakoidov). Chlorofyl a, modrozelený, aktívnejší, prevyšuje obsah žltozeleného chlorofylu b 2–3-násobne u tienistých, 5 až viacnásobne u slnečných listov. V sušine listov tvoria chlorofyly asi 0,6–1,2 % podľa rastlinného druhu, polohy v rámci koruny, fázy vývinu, podmienok osvetlenia, výživy a iné. Denná obnova chlorofylov činí 5 až 8 % z celkového obsahu. Porfyrínové časti chlorofylových molekúl sú viazané na nepolárne, lineárne reťazce fytolu, ktoré ich zakotvujú v membráne chloroplastu, čím zabezpečujú ich určitú polohu.
2. karotenoidy: oranžový karotén (α- a β-karotén) a žlté až hnedé xantofyly (xantofyl, zeaxantín, violaxantín a iné). Karotenoidy sú deriváty izoprénu, avšak sú usporiadané lineárne. Sú (ako u chlorofylov) typické striedaním dvojných a jednoduchých väzieb s delokalizovanými, voľnými π-elektrónmi.
Vďaka tomu sú oba systémy schopné prijímať len žiarenie o určitom rozsahu vlnovej dĺžky (t. j. v „oknách“ spektra), ostatné žiarenie odrážajú či prepúšťajú späť. Toto odrazené žiarenie vníma naše oko ako doplnkovú farbu k farbe pohlcovanej (napr. listy sa nám javia ako zelené, pričom pohlcujú žiarenie v modrej a červenej oblasti viditeľného spektra). Karotenoidy majú funkciu svetlozberateľskú, antioxidačnú a ekologickú (upozorňovací signál pre opeľovačov a pre živočíchy, ktoré roznášajú semená pozretím farebných plodov).
Prvým krokom fotosyntézy je zachytenie žiarivej energie. Na vyšších úrovniach ide o prienik žiarenia, ovplyvňovaný priestorovým usporiadaním vetiev a listovej plochy v rámci koruny stromu (či nadzemného systému kra, liany a iných foriem), ďalej o usporiadanie chlorenchýmu (všeobecne pletív s bunkami obsahujúcimi chloroplasty), usporiadanie chloroplastov vnútri buniek a nakoniec o usporiadanie pigmentových svetlozberateľských (anténnych) sústav.
Vlastným reakčným centrom fotosyntézy je chlorofyl a. Ostatné farbivá slúžia ako doplnkové farbivá, ktoré okolo reakčného centra tvoria spolu s enzýmami a bielkovinami tzv. svetlozberateľské antény – tie slúžia na príjem fotónov a usmerňovanie ich toku na chlorofyl a.
Primárne a sekundárne procesy fotosyntézy krok za krokom
V prvej fáze (primárne procesy fotosyntézy) je energia zo slnečného žiarenia zachytená a prechodne uskladnená do energie chemických väzieb špeciálnych malých molekúl adenozíntrifosfátu – ATP (táto molekula všeobecne u všetkých živých, aj živočíšnych, buniek slúži ako zásobáreň okamžite využiteľnej energie). Súčasne je energia uložená do molekuly redukčného činidla – koenzýmu nikotínamidadeníndínukleotidfosfátu, skrátene označovaného ako NADPH. Obe molekuly sú tzv. primárnymi produktmi fotosyntézy a slúžia ako prenášače energie a reaktívnych chemických skupín. Procesy tejto fázy sa uskutočňujú na membránach tylakoidov chloroplastov v dvoch vzájomne spriahnutých, na seba nadväzujúcich krokoch pomocou fotosystémov (komplexov molekúl bielkovín a stoviek molekúl fotosyntetických pigmentov). Ide o fotosystém I, vývojovo starší, vyskytujúci sa už u fotosyntetizujúcich baktérií, a fotosystém II, vývojovo mladší.

Fotosystém I absorbuje žiarivú energiu o vlnových dĺžkach okolo 700 nm (reakčné centrum P-700) a podieľa sa na generovaní ATP; fotosystém II pohlcuje žiarenie o kratších vlnových dĺžkach (reakčné centrum P-680), rozkladá molekulu vody za vzniku molekulárneho kyslíka a zachytáva vodík pre redukciu NADP. Kyslík je ako vedľajší produkt primárnych procesov uvoľňovaný do ovzdušia a je nevyhnutný pre dýchanie všetkých organizmov typu aeróbov (vrátane človeka).
V druhej fáze (sekundárne procesy fotosyntézy) sú využívané molekuly primárnych procesov, ktoré dodávajú energiu a redukčný potenciál – tj. „poháňajú" proces fixácie uhlíka. Pri ňom sú z plynného atmosférického oxidu uhličitého (CO₂, ktorý do rastliny vstupuje prieduchmi či lenticelami) a z vody (H₂O, ktorú prijímajú rastliny svojimi koreňmi z pôdy) vyrábané cukry. Dej sa odohráva v stróme chloroplastov. Nepotrebuje svetlo, je však závislý na primárnej fáze – krátko po zotmení prestane prebiehať tiež.
Primárne procesy fotosyntézy možno súhrnne vyjadriť vstupnými faktormi a výstupnými produktmi nasledovne: žiarivá energia (8 fotónov) + chlorofyly + 2 NADP⁺ + 2 ADP + fosfát + 2 H₂O → 2 NADPH₂ + 2 ATP + O₂
Sekundárne procesy využívajú produktov z procesov primárnych a obnovujú vstupné faktory: nCO₂ + nAcc + 2n NADPH₂ + 2n ATP → glukóza + 2n NADP⁺ + 2n ADP + 2n fosfát + nAcc + tepelná energia
Molekuly sacharidov a molekuly ATP (s energeticky bohatou fosfátovou väzbou, vzniknutou fotofosforylovaním) sa potom použijú ako zdroje energie chemických väzieb a zdroje materiálu pre prípravu mnohých iných malých organických molekúl, ktoré sú pre rastlinnú bunku nepostrádateľné.

C3 a C4 fotosyntéza: Calvínov cyklus a Hatch-Slackova dráha
Proces začlenenia uhlíka z CO₂ do organických väzieb závisí od druhu rastliny a u drevín môže prebiehať dvoma typmi. V Calvinovej ceste je akceptorom CO₂ päťuhlíkový sacharid (ribulóza-1,5-bisfosfát), ktorý sa po prijatí atómu uhlíka rozpadá na dve trojuhlíkové molekuly. Túto cestu realizujú druhy nazývané C₃ rastliny a karboxyláciu katalyzuje špecifický enzým Rubisco (ribulóza-1,5-bisfosfát karboxyláza). V Hatch-Slackovej ceste je akceptorom CO₂ trojuhlíkový akceptor (fosfoenopyruvát), ktorý po zabudovaní atómu uhlíka poskytuje priamo štyruhlíkový produkt. Ide o typ fotosyntézy u tzv. C₄ rastlín, u ktorých sa uplatňuje ako enzým Rubisco, tak enzým PEP (fosfoenolpyruvát karboxyláza). Konečnými produktmi oboch syntetických ciest sú sacharidy (fotosyntáty) – východiskové zlúčeniny všetkých ostatných látok tela – a obnovený príjemca CO₂ (t. j. päťuhlíkový alebo trojuhlíkový akceptor – Acc).
V našich podmienkach prevažujú rastliny s C₃ fotosyntézou. Sú citlivejšie na vyššie teploty, suchá a nedostatok dusíka. Hatch-Slackov cyklus prebieha u radu teplomilných druhov rastlín, napr. tropických. Pre rastliny s týmto cyklom sú typické pošvy okolo cievnych zväzkov s bunkami obsahujúcimi chloroplasty. Existujú však aj také prípady, keď na jednej rastline nájdeme listy s C₃ fotosyntézou aj listy s C₄ typom (u nás napr. u kukurice).
Čo ovplyvňuje intenzitu fotosyntézy: svetlo, voda, teplota aj minerály
Vplyv vonkajších a vnútorných činiteľov na fotosyntézu

Konečnými produktmi fotosyntézy sú sacharidy (fotosyntáty), východiskové zlúčeniny všetkých ostatných látok tela a obnovený prijímač (akceptor) CO₂. Úroveň fotosyntézy, vzhľadom na ostatné životné procesy (respiráciu), sa prejavuje spotrebou CO₂ (resp. výdajom O₂, čistá fotosyntéza). Kapacitou fotosyntézy druhu (v danej fáze vývinu a aktivity) sa rozumie najvyššia rýchlosť príjmu CO₂ pri prirodzenom obsahu CO₂ v atmosfére a optimálnych podmienkach ostatných faktorov (svetla, vody, teploty, živín a pod.). Potenciálna čistá fotosyntéza sa stanovuje navyše aj za optimálneho tlaku CO₂. Kompenzačný bod (vyjadrovaný ožiarenosťou, dostupnosťou CO₂, teplotou a pod.) je stav dreviny, resp. jej časti – vetvy, listov, s nulovou výmenou CO₂, t. j. vyrovnanou rýchlosťou fotosyntézy (spotrebou) a respirácie (výdajom CO₂). Ak podmienky kompenzačného bodu trvajú dlhodobo, rastlina, alebo jej časť, chradne a predčasne odumiera. U drevín v zatienení ide často o obmedzený rast, zníženú odolnosť voči patogénom a tiež stratu bazálnej časti koruny (tzv. samočistenie koruny, t. j. odumieranie bazálnych, silne zatienených vetiev). Teplotné nároky na fotosyntézu sú trochu nižšie než na respiráciu. Teplotné optimum pre naše domáce, stromovito rastúce druhy drevín je okolo 25 °C. Ihličie ihličnanov môže vykonávať primárne deje fotosyntézy aj pri teplotách okolo 0 °C. Pri mrazových teplotách sa rozpadá fotosystém II.
Pôsobením vysokých dávok slnečného žiarenia (pri dlhom dni, za jasného počasia, okolo poludňajších hodín) môže byť odštiepený tetrapyrol od fytolu (defytolácia chlorofylu), a tým sa chlorofyl stáva neúčinným – poškodená časť listu je vybielená. Pôsobením kyslého prostredia je v molekule chlorofylu atóm horčíka nahradený atómom vodíka (chlorofyl prechádza na feofytín, proces sa označuje ako feofytinácia). Aj tým sa stáva chlorofyl vo fotosyntéze neúčinným, jeho zelená farba sa mení na žltohnedú až hnedú (účinok „kyslých dažďov", hnednutie opadaného lístia pri jeho rozklade).
Najčastejšie býva fotosyntéza znižovaná nedostatkom vody, zníženou transpiráciou, a tým zvýšeným ohrevom listov. S nedostatočným transpiračným tokom môže byť fotosyntéza obmedzená tiež suboptimálnou minerálnou výživou, a to radom živín, ktoré sú pre tento dej nevyhnutné (napr. N, P, Fe, Mg, Cu, K, Mn, Cl). Navyše dochádza pri nedostatku vody v listoch k uzavretiu prieduchov a k zvýšeniu odporu listovej pokožky proti vstupu CO₂ do listu. Pri dlhšom vodnom strese sa neodvádzajú z listov fotosyntáty, hromadia sa v nich a fotosyntéza ustáva. Rýchlosť fotosyntézy závisí teda v značnej miere od rýchlosti difúzie plynov (CO₂, O₂, vodných pár), čo je dané nielen vonkajšími faktormi, ale najmä tým, či je drevina súčasťou porastu alebo rastie solitérne, aký má tvar a usporiadanie koruny a listov, aký je povrch listov, funkcia ich prieduchov, usporiadanie mezofylu, systému vodivých pletív a pod.
Zelené rastliny obsahujú vo svojich bunkách ďalšie systémy molekúl, ktoré sú schopné využívať energiu slnečného žiarenia. Ich činnosť je však mnohostrannejšia než činnosť chlorofylov. Ide o fytochróm – systém dvoch foriem jedného typu molekuly, ktorá je zložená z chromoforu a bielkovinovej časti. Formy P₆₆₀ a P₇₃₀ medzi sebou prechádzajú po pohlcení fotónu o zodpovedajúcej vlnovej dĺžke a sú fotoreceptorom rastlinných buniek v červenej oblasti svetla. Prostredníctvom fytochrómu reguluje svetlo aktivitu radu enzýmov, syntézu nukleových kyselín, bielkovín, chlorofylov a pod., klíčenie semien, kvitnutie rastlín, vývoj a opadávanie listov, utváranie pukov a pod.

Výmena CO₂ a kyslíka u rastlín: bilancia uhlíka a kompenzačný bod
Výmena CO₂ u rastlín
Bunkový metabolizmus uhlíka spája s vonkajším prostredím výmena plynov. Pri fotosyntéze spotrebúvajú chloroplasty CO₂, ktorého prísun musí byť zabezpečený, a uvoľňujú kyslík. Súčasne bunky vo dne aj v noci prijímajú kyslík, potrebný na dýchanie, a uvoľňujú oxid uhličitý. Vo fotosyntetizujúcich listoch môže v danom čase prevládať jeden alebo druhý z týchto protichodných procesov. Dýchanie, prebiehajúce na svetle, sa skladá z fotorespirácie a mitochondriálneho dýchania. V priebehu svetlej časti dňa je rýchlosť príjmu CO₂, potrebného pre fotosyntézu, na jednotku biomasy spravidla väčšia ako rýchlosť uvoľňovania CO₂ celkovým dýchaním na svetle; výsledkom je čistý príjem CO₂ listom. Ak rýchlosť fotosyntézy klesá, môže sa stať, že súčasne prebiehajúce dýchanie sa jej práve vyrovná (je dosiahnutý kompenzačný bod, pozri vyššie). Klesá-li rýchlosť fotosyntetickej aktivity ešte ďalej, dýchanie prevládne a za tmy prebieha iba respiračné uvoľňovanie CO₂. Vzniká stav zápornej bilancie uhlíka, ktorá dlhodobo znamená zánik jedinca.
Metabolizmus na hranici kompenzačného bodu majú napr. zatienené listy na báze koruny alebo na vnútornom okraji tzv. korunového jadra. Ide o bezlistý priestor, v ktorom nie sú dostatočné podmienky pre dlhodobú kladnú bilanciu uhlíka. Kompenzačný bod môže byť spôsobený aj vyššou teplotou listov (tj. nad 35 °C), ktorá urýchľuje dýchanie lístia a zároveň spomaľuje fotosyntézu – podobne môže pôsobiť rad stresových faktorov vrátane obranných reakcií po poranení a pod.
Vzťah fotosyntézy a respirácie: ako rastliny uzatvárajú uhlíkový cyklus
Fotosyntéza a respirácia

Fotosyntéza a respirácia sú komplementárne (vzájomne sa dopĺňajúce) deje, a to nielen vnútri rastlinných buniek, t. j. buniek listového mezofylu, kde oba deje prebiehajú súčasne v oddelených kompartmentoch chloroplastov a mitochondrií. Na inej úrovni prebiehajú oba deje v rámci jednej rastliny, kde fotosyntéza aj dýchanie prebiehajú v jej zelených častiach (listoch, stopkách, mladých steblách a pod.), zatiaľ čo iba respirácia prebieha v živých bunkách, ku ktorým nepreniká dostatok svetla (bunky vo vnútri konárov, kmeňov, bunky koreňov). Potom napr. dýchanie buniek kambiálnej zóny prispieva k dodávaniu oxidu uhličitého pre fotosyntézu buniek primárnej kôry. V širšom meradle môžeme o vzťahu medzi fotosyntézou a respiráciou uvažovať ako o vzťahu medzi producentmi (zelenými rastlinami) a reducentmi a konzumentmi (t. j. heterotrofnými organizmami), zdanlivo úplne oddelenými od rastlín. To znamená, že nie všetky výmeny medzi rastlinami a živočíchmi sú jednosmerné. Rastliny, živočíchy a baktérie na tejto planéte existovali vedľa seba tak dlho, že niektoré z týchto organizmov sa stali prirodzenou súčasťou životného alebo aj vnútorného prostredia iných organizmov (lišajníky – komplexné organizmy heterotrofnej huby a autotrofnej sinice). A niektoré molekuly CO₂, ktoré sú dnes fixované do organických molekúl pri fotosyntéze v zelenom liste, boli včera uvoľnené do atmosféry pri dýchaní živočícha alebo baktérie rozkladajúcej mŕtvu organickú látku. Vidíme teda, že využitie uhlíka tvorí cyklus, ktorý zahŕňa biosféru (všetky živé organizmy na zemi) ako celok a prekračuje hranice medzi jednotlivými organizmami. Podobne sa pohybujú medzi živým a neživým svetom atómy dusíka, fosforu a síry v cykloch, ktoré postihujú rastliny, živočíchy, huby a baktérie.

Dýchanie drevín: glykolýza, Krebsov cyklus a oxidatívna fosforylácia
Dýchanie (respirácia)
Bunky získavajú energiu oxidáciou organických molekúl. Zemská atmosféra obsahuje veľký podiel kyslíka, v prítomnosti ktorého je energeticky najstabilnejšou formou uhlíka CO₂ a najstabilnejšou formou vodíka H₂O. Bunka je preto schopná získavať energiu zo sacharidov či iných organických molekúl tak, že umožní zlučovanie ich atómov uhlíka a vodíka s kyslíkom za vzniku CO₂ a H₂O. Uvoľnenú energiu ukladá do makroergických väzieb ATP. Tento dej sa nazýva respirácia (dýchanie) a získavanie energetickej „konzervy“ oxidatívna fosforylácia.
Dýchajú všetky organizmy, resp. všetky ich živé bunky. (Iným mechanizmom rozkladu organickej hmoty je rýchla oxidácia pri spaľovaní, napr. dreva, ropy, zemného plynu, rašeliny a pod.).
Súhrnná chemická rovnica dýchania sa niekedy zjednodušuje a vyjadruje sa ako opak fotosyntézy, tj. ako oxidácia glukózy za uvoľnenia energie, oxidu uhličitého a vody:
V skutočnosti je dýchanie proces oveľa zložitejší, naviazaný na produkty fotosyntézy, prebieha riadene cez rad postupných krokov a vychádza z neho sieť syntetických dráh a koordinácií. Preto je lepšie chápať respiráciu ako metabolické centrum. Dýchaním sa v rastline rozkladajú zložité látky s vysokou potenciálnou energiou (napr. cukry, tuky, bielkoviny) na látky jednoduché a pri tom sa uvoľňuje energia. Získaná energia slúži pre rôzne funkcie bunky – vnútrobunkový pohyb, udržiavanie a obnovovanie štruktúr, syntézy, uvoľňovanie tepla (napr. na jar v napučaných púčikoch a nakvetlých púčikoch). Rozklad respirovaného substrátu (napr. glukózy) prebieha v mnohých postupných krokoch: glykolýza, dekarboxylácia pyruvátu, cyklus kyseliny citrónovej a oxidácia konečného akceptora elektrónov (NADH₂). Glykolýza je lokalizovaná v cytoplazme, zatiaľ čo cyklus kyseliny citrónovej a oxidatívna fosforylácia prebiehajú v mitochondriách. Výťažkom celého rozkladu je 36 molov ATP (adenozíntrifosfát), 2 moly GTP (glyceraldehydtrifosfát) a voľná entalpia ΔG = -2,87 MJ na 1 mol glukózy. Okrem uvoľňovania energie vznikajú pri postupných rozkladných krokoch aj veľmi potrebné medziprodukty, ktoré sú východiskovými prekurzormi pre následné syntézy (napr. lignínu).
Respirácia prebieha s vyššou účinnosťou za prítomnosti kyslíka (aeróbne dýchanie), ale môže sa diať aj bez molekulárneho O₂, pomocou reťazca oxidoredukčných reakcií, ústiacich do molekúl jednoduchších, ale stále ešte bohatých na energiu. Takejto respirácii hovoríme anaeróbna a rozlišujeme ju na kvasenie (rozklad sacharidov a lipidov) a hnitie (rozklad organických látok obsahujúcich dusík). Kvasenie sa delí podľa konečného, hromadeného a ďalej nespracovávaného produktu napr. na kvasenie etanolové (lihové), mliečne, maslové a pod. Týmito procesmi získavajú energiu prevažne mikroorganizmy (baktérie, kvasinky a pod.).
Proces dýchania – oxidácia glukózy
Dýchanie prebieha v dvoch po sebe nasledujúcich krokoch – začína anaeróbnou glykolýzou a pokračuje aeróbnym štiepením kyseliny pyrohroznovej alebo kvasením.
Anaeróbna glykolýza (anaeróbne štiepenie glukózy)
Prebieha (v cytoplazme) za katalytického pôsobenia enzýmov v nej rozpustených a vždy za neprístupu kyslíka, a to aj v bunkách, ktoré sa vyskytujú v aeróbnom prostredí. Tieto dva fakty svedčia o starobylosti tejto metabolickej dráhy. Konečným produktom sú dve molekuly kyseliny pyrohroznovej (pyruvát) a ATP.
Aeróbne štiepenie kyseliny pyrohroznovej (v mitochondriách)
Rozlišujeme tri časti priebehu – dekarboxyláciu kyseliny pyrohroznovej, Krebsov cyklus a dýchací reťazec: – dekarboxylácia kyseliny pyrohroznovej: Z molekuly kyseliny pyrohroznovej (CH₃COCOOH) sa odštiepi jeden atóm uhlíka vo forme CO₂ (kyselina je dekarboxylovaná) a za pôsobenia acetylkoenzymu A vzniká aktivovaná kyselina octová. – Krebsov cyklus (cyklus kyseliny citrónovej, citrátový cyklus): Prebieha v matrix. Aktivovaná kyselina octová reaguje s kyselinou oxaloctovou a vzniká kyselina citrónová. Uvoľňuje sa molekula ATP a vodík. Tri molekuly koenzymu NAD⁺ a jedna molekula koenzymu FAD (flavínadeníndínukleotid) sa naviazaním vodíka redukujú a prenášajú ho do dýchacieho reťazca. – dýchací reťazec: Je to reťaz oxidačno-redukčných reakcií. Protóny aj elektróny prinášajú redukované koenzýmy NADPH + H⁺ a FADH₂. Protóny sa na začiatku uvoľňujú do prostredia. Elektróny sú prenášané radom prenášačov vo vnútornej membráne mitochondrií po spáde elektrického potenciálu a pri tom sa syntetizuje ATP. Syntézu ATP v dýchacom reťazci označujeme ako oxidatívnu fosforyláciu. Je veľmi podobná fosforylácii, prebiehajúcej na tylakoidoch počas fotosyntézy. Pri prenášaní elektrónov sa uvoľňuje energia, vďaka ktorej je umožnený presun protónov cez membránu z matrix do medzimembránového priestoru, kde sa zvyšuje ich koncentrácia. Tak vzniká na mitochondriálnej membráne protónový gradient (gradient pH), spôsobený rozdielnou koncentráciou protónov v matrix a medzimembránovom priestore. Protóny sa späť do matrix vracajú cez enzým ATP-áza, ktorá energiu uvoľnenú pri prechode protónov ukladá do ATP. Na syntézu 1 molekuly ATP je potrebný prechod dvoch protónov do matrix. Pri tejto fosforylácii vzniká viac ATP ako počas fotosyntézy. Na konci reťazca sa elektróny prenášajú na molekulárny kyslík, a tak vznikajú kyslíkové anióny, ktoré sa zlučujú s protónmi na vodu.
2 H⁺ + O₂⁻ = H₂O
Pri oxidačných pochodoch v mitochondriách vznikne celkovo 36 molekúl ATP z jednej molekuly glukózy.
Enzýmy, ktoré katalyzujú reakcie Krebsovho cyklu a dýchacieho reťazca, sa nachádzajú a vznikajú vo vnútornej membráne mitochondrií.
Kvasenie (fermentácia)
Kyselina pyrohroznovú je odbúravaná bez prístupu kyslíka a reakcie sa zastavujú na medziproduktu – napr. etanole. V takom prípade ide o etanolové kvasenie, ktoré môže prebiehať aj v rastlinách za neprístupu vzduchu (pozri vyššie). Etanol je však toxický, najmä pre prácu mitochondrií; pri dlhotrvajúcich anaeróbnych podmienkach (dostavujúcich sa najmä v lete) môžu dreviny odumierať. Odolnejšie sú dreviny luhov s pravidelnými jarnými záplavami alebo stromy vo vyšších polohách, rastúce v blízkosti bystríc – rýchlo tečúca a chladná voda je dostatočne okysličená. Kvasením sa v porovnaní s aeróbnym dýchaním uvoľňuje menej energie, pretože väčšina zostáva viazaná v konečných produktoch.
Čo ovplyvňuje rýchlosť dýchania drevín v priebehu roka
Aj u drevín môžu nastať nepriaznivé podmienky pre aeróbnu respiráciu koreňov (zaplavenie, zhutnenie pôdy). Navyše, pri zníženej aerácii pôdy, produkujú anaeróbne pôdne baktérie potenciálne toxické zlúčeniny vrátane plynov, organických zlúčenín – alkoholov, karbonylových zlúčenín, volatilných mastných kyselín, nevolatilných kyselín, fenolických kyselín a volatilných zlúčenín síry. Vtedy býva ovplyvnený aj rast výhonkov stromov obmedzením predlžovania ich internódií a obmedzením tvorby a plošného rastu listov, indukciou starnutia listov, ich poškodením a opadom. Pri dlhodobom vplyve sa znižuje hrúbkový rast vzrastlých stromov a tiež sa mení anatómia floému aj xylému. Ďalej je drasticky redukovaná tvorba a rast koreňov a urýchľovaný ich úbytok v dôsledku aktivity plesní rodu Phytophthora. Znížená respirácia koreňov má vplyv na zníženie rýchlosti fotosyntézy, pretože navodzuje uzatváranie prieduchov a mení karboxylačné enzýmy, znižuje obsah chlorofylu.

Absorpcia minerálnych živín klesá so zmenšeným rozsahom koreňového systému a tiež s nedostatkom energie, ktorej sa pri anaeróbnej respirácii uvoľňuje menšie množstvo, nedostačujúce pre všetky funkcie koreňov (môže dôjsť aj k vyplavovaniu iónov z koreňov do pôdy). Modifikácia a redukcia koreňového rastu robí stromy náchylnými na vyvrátenie. Hladina auxínu, etylénu a kyseliny abscisovej v stonke rastie, zatiaľ čo hladina giberelínov a cytokininov klesá. Celkovo oslabené dreviny ľahšie podliehajú napadnutiu biotickými činiteľmi.
Rastliny, resp. ich živé bunky a najmä meristémy, dýchajú celý deň aj noc, fotosyntéza prebieha iba cez deň (v svetlej časti dňa). V listoch niekoľkonásobne prevyšuje dýchanie (asi päť- až desaťnásobne). Pri respirácii je najrýchlejšie a najlepšie využiteľná energia z jednoduchých sacharidov, ktoré sa v telách organizmov nemôžu skladovať vo vyšších koncentráciách, pretože sú osmoticky aktívne. Spravidla sa premieňajú na iné formy zásobných látok – polysacharidy (najčastejšie škrob), tuky (najmä zásobné v semenách, napr. Juglans, Fagus, Corylus), niekedy aj zásobné bielkoviny. Stavebné látky bunkových stien, hoci by boli cukrovej povahy, ako je celulóza, hemicelulózy, pektáty a pod., dreviny spätne ako zdroj energie využiť nemôžu.
Napokon vydýchateľným substrátom sú aj bielkoviny, ktorých stavebné kamene – aminokyseliny – môžu byť znovu využité pre nové alebo iné, lepšie zodpovedajúce štruktúry (napr. pri prestavbe bunkových membrán).
Faktory ovplyvňujúce dýchanie
U jednotlivých rastlín dýchajú kvety a nezrelé plody väčšou rýchlosťou než listy a korene dýchajú rýchlejšie než stonkové časti výhonkov. U vetiev a kmeňov stromov sú hlavnými oblasťami dýchania kôra – najmä felogén, živá časť lyka, kambiálna zóna a vonkajšie, funkčné vrstvy dreva, tj. biel s obsahom živých buniek. Rastové dýchanie (dýchanie vo vegetačnom období, pri raste a syntetických procesoch buniek) je u semenáčov, koreňových a stonkových vrcholov, pri rozvádzaní listov a vývoji plodov 3 až 10-krát vyššie než dýchanie udržiavacie (napr. zimné dýchanie ihličia). Rýchlosť dýchania je priamo úmerná rýchlosti rastu. S pokračujúcou diferenciáciou a dozrievaním pletív sa respiračná aktivita vracia na oveľa nižšiu úroveň. Prechodné, ostro zvýšené dýchanie sprevádza zrenie plodov a starnutie listov (tzv. klimakterické dýchanie). Ide o štiepenie energeticky bohatých látok, sťahovanie pohyblivých minerálnych živín (napr. draslíka, dusíka), zároveň o ukladanie splodín metabolizmu do orgánu, ktorý bude od rastliny oddelený a o prípravu jeho odlučovacej, resp. hojivej vrstvy. Tieto enzymatické procesy prirodzeného starnutia sú prísne geneticky riadené a závislé od priaznivých podmienok prostredia. Prejavujú sa napr. postupným jesenným sfarbovaním listov (keď bunky okolo žilnatiny sú najdlhšie živé) – na rozdiel od náhleho odumretia (napr. suchom) za zelena a za oneskoreného opadu. Odumieranie rastlinných buniek je často sprevádzané uvoľňovaním plynných metabolických produktov (napr. etylénu).
Napr. v zime sa bunky vetiev jaseňa udržujú pri živote a respirácia klesá asi na 2 mg CO₂ za hodinu na 100 g čerstvej hmotnosti vetvy (prípadne aj menej). V máji, pri teplote 18,5 °C, stúpa rýchlosť respirácie s rastom dreviny a vrcholí v júni až júli, keď sa uvoľňuje až 6 mg CO₂ za hodinu na 100 g čerstvej hmotnosti biomasy. Akonáhle v septembri rast ustáva, klesá aj dýchanie. U drevín stúpa respirácia pri sekundárnom hrubnutí a udržuje sa dosť vysoko až do konca augusta, u duba a buka klesá na konci októbra, u lipy až v decembri. V zime dýchanie takmer nekolíše a zotrváva na úrovni o ¹/₃ až ¹/₄ nižšej než za maxima v lete. Rýchlosť dýchania sa teda mení nielen počas dňa, ale aj v priebehu roka. Jednotlivé druhy rastlín za rovnakých podmienok vykazujú rôznu rýchlosť dýchania. Napríklad listy opadavých stromov dýchajú priemerne päťkrát väčšou rýchlosťou než asimilačné orgány stálezelených stromov. Zvyčajne býva pomer uvoľneného CO₂ k prijatému kyslíku (respiračný koeficient) rovný jednej. Platí to ovšem vtedy, keď je pri dýchaní spaľovaný škrob; vydychuje-li sa tuk (olejnaté semená – Fagus, Juglans), klesá hodnota respiračného koeficientu pod jednu. Avšak z dlhodobého hľadiska prežitia rastliny musí byť celková ročná bilancia medzi fotosyntézou a dýchaním kladná, tj. fotosyntéza musí nad respiráciou prevládať, inak by nemohol byť zabezpečený rast, tj. zväčšovanie a rozrastanie biomasy.
Rýchlosť dýchania je závislá od vnútorných aj vonkajších faktorov. Z vnútorných faktorov (okrem druhových a individuálnych odchýlok) majú vplyv na intenzitu dýchania: 1. rast, 2. dostupný substrát, tj. koncentrácia organických zlúčenín v bunkách (s vyššou koncentráciou stúpa aj intenzita dýchania), 3. množstvo vody v pletivách.
Semená intenzívnejšie dýchajú, akonáhle nabobtnajú vodou. Z vonkajších činiteľov dýchanie ovplyvňuje najmä: 1. teplota: pri zvýšenej teplote (až do 40 °C) dýchanie stúpa, pri nižších teplotách rýchlosť dýchania klesá. Ihličie borovice pri 0 °C dýcha 25-krát rýchlejšie než pri -12 °C, medzi 0 až 40 °C stúpa rýchlosť dýchania 2 až 3-krát na každých 10 °C, pri teplotách medzi 50 °C a 60 °C sú enzýmy a funkčne dôležité membránové štruktúry teplom poškodzované a dýchanie ustáva, 2. svetlo celkovo nemá pri dýchaní význam, 3. vlhkosť vzduchu pôsobí priaznivo, 4. vyššia koncentrácia CO₂ v ovzduší dýchanie spomaľuje, až pôsobí škodlivo, 5. pri poranení sa zvyšuje intenzita dýchania (súvisí s aktívnou prípravou a delením buniek).
Pre normálny priebeh mitochondriálneho dýchania postačuje koncentrácia kyslíka 1 až 3 %. Keď koncentrácia klesne pod túto hladinu, poklesne najprv rýchlosť dýchania, príjem vody a iónov a rast koreňov. Zvyšuje sa produkcia kyseliny abscisovej a najmä etylénu, takže nadzemný rast je narušený a listy predčasne opadávajú. U vyšších rastlín môže za nepriaznivých okolností dôjsť aj k dýchaniu bez kyslíka (anaeróbne dýchanie – viď vyššie). Môže sa tak stať, ak sú dreviny (ich korene) zaplavené vodou, alebo ak klíči semeno v zabahnenej pôde. Je to však iba východisko z núdze, a to iba na krátky čas. Pri dlhodobejšom nedostatku vzduchu rastliny odumrú. Rastliny rastúce v obzvlášť ťažkých pôdach, kde kyslík difunduje pomaly, vyvíjajú extenzívny povrchový koreňový systém a adventívne korene. Extrémnu špecializáciu predstavujú dýchacie korene. Dreviny vydychujú takmer vždy cukry. Ak potrebujú k dýchaniu iné látky, napr. tuky, premieňajú ich na cukry. Iba výnimočne spotrebúva rastlina bielkoviny.

Fotorespirácia: svetelné dýchanie a jeho vplyv na produktivitu stromov
Fotorespirácia
V rastlinných častiach, ktoré obsahujú chlorofyl, prebieha v spojení s fotosyntézou metabolický proces podobný dýchaniu, pri ktorom sa na svetle spotrebúva O₂ a uvoľňuje CO₂, ale ktorý – na rozdiel od dýchania – v tme prestáva. Táto výmena O₂ za CO₂ bola nazvaná svetelnou respiráciou alebo fotorespiráciou. Prebieha spočiatku v chloroplastoch, prechádza do peroxizómov a ukončuje sa v mitochondriách. Enzýmom fotorespiračného metabolizmu je Rubisco (ribulóza-1,5-bisfosfát karboxyláza-oxygenáza). Pri príjme kyslíka štiepi ribulózu-1,5-bisfosfát, pričom sa uvoľňuje už raz prijatý CO₂. Pomer medzi fotorespiráciou a fotosyntézou je regulovaný prísunom O₂ a CO₂, hustotou osvetlenia, teplotou a pod. Vysoký parciálny tlak O₂ podporuje fotorespiráciu, veľký prísun CO₂ podporuje fotosyntézu.
Za letných vonkajších podmienok (21 % O₂ a 0,03 % CO₂ vo vzduchu, silné osvetlenie, teplota medzi 20 až 30 °C) strácajú dreviny okolo 20 %, alebo v extrémnom prípade až 50 % (smrekové ihličie), fotosynteticky získaného CO₂ vo forme fotorespiračného CO₂. Na druhú stranu, tento proces spotrebúva energiu (obmedzuje ohrev listov) a poskytuje prekurzory pre obnovu chlorofylu.
Voda v živote drevín: fyzikálne vlastnosti a ich biologický význam
Vodný režim drevín

Všetky životné formy na našej planéte sú funkčne priamo závislé od vlastností vody a od látok, ktoré sa vo vode rozpúšťajú (ióny anorganických solí, jednoduché sacharidy, bielkoviny, organické kyseliny a i.), alebo sú zmáčavé, hygroskopické a napučiavacie (celulóza, drevo, koloidy), či naopak sú vodoodpudivé (látky povahy tukov, lipidov). Z fyzikálnych vlastností vody sú najvýznamnejšie tieto: – merné skupenské teplo topenia ľadu (množstvo energie potrebnej na prechod jednotkového množstva látky z pevnej fázy do fázy kvapalnej) je 334 kJ . kg⁻¹, – merná tepelná kapacita (množstvo energie potrebnej na ohriatie jednotkového množstva látky o 1 °C) vody v kvapalnom stave je 4 186 kJ . kg⁻¹. K⁻¹, – merné skupenské teplo varu (množstvo energie potrebnej na prechod jednotkového množstva látky z kvapalnej fázy do plynnej) je 2 260 kJ . kg⁻¹,
čo sú hodnoty v porovnaní s inými látkami mimoriadne vysoké. Pri premene skupenstva kvapalného na pevné dochádza k zväčšeniu objemu, priamym dôsledkom je zníženie hustoty (t. j. ľad pláva na vode). Približne platí, že hustota vody je 1 000 kg . m⁻³, t. j. 1 l vody váži 1 kg.
Teplotná objemová rozťažnosť je u oboch skupenstiev (kvapalného aj pevného) úplne zanedbateľná v porovnaní s objemovými zmenami pri zmene skupenstva, topení či tuhnutí. Možno teda povedať, že voda v kvapalnej ani pevnej fáze svoj objem s teplotou prakticky nemení. Nepatrná závislosť je však merateľná a je známe, že voda má najvyššiu hustotu pri 4 °C.
Ako pri každej kvapaline, tak aj u vody pozorujeme povrchové napätie. Ide o energiu, ktorú je potrebné vynaložiť na to, aby sa povrch kvapaliny zväčšil o 1 m². Táto hodnota dosahuje 0,072 J . m⁻² (napr. v stave beztiaže zaujíma voda tvar gule, ktorá má najmenšiu plochu vzhľadom k objemu). Viskozita, zjednodušene povedané, opisuje, ako ľahko kvapalina tečie. Čím je nižšia viskozita (vnútorné trenie), tým menší odpor kvapalina kladie, napr. pri prietoku trubicou (v prípade drevín cievou, cievičkou).
Elektrická vodivosť čistej vody je za normálnych podmienok taká nízka, že ju môžeme považovať za veľmi dobrý izolant. Zanedbateľnú elektrickú vodivosť spôsobuje nepatrné množstvo disociovaných molekúl (prítomných vo forme oxóniových iónov H₃O⁺ a hydroxylových iónov OH⁻). Avšak rozpustením ďalších, ľahko disociujúcich látok sa elektrická vodivosť vodného roztoku môže zvýšiť až o niekoľko rádov.
Molekula vody obsahuje dva atómy vodíka, ktoré s centrálnym atómom kyslíka zvierajú uhol 105° (v pevnej fáze – ľade); v kvapalnej fáze uhol okolo tejto hodnoty mierne osciluje. Hoci navonok sa molekula vody správa ako neutrálna, protóny umiestnené od povrchu atómu kyslíka v uvedenom uhle spôsobujú jej polaritu, t. j. slabý kladný náboj na jednej jej strane. Výsledkom je, že kladne nabitá strana jednej molekuly vody je priťahovaná k negatívnej strane susednej molekuly vody alebo molekuly inej. Tieto slabé väzby sa označujú ako vodíkové a existujú aj medzi radom organických molekúl (s energiou od 8 do 42 kJ . mol⁻¹). V prípade vody zodpovedajú za jej anomálne správanie – je kvapalná v rozmedzí 0 až 100 °C, je polárnym rozpúšťadlom, tvorí pravé aj koloidné roztoky. Súdržnosť molekúl vody navzájom sa označuje ako kohézia, súdržnosť molekúl vody s molekulami iného typu je adhézia. Pomocou kohézie a adhézie vody a pomocou vodíkových väzieb medzi radom iných štruktúr je zabezpečená súdržnosť rastlinných tiel.
Konštantný objem a nestlačiteľnosť kvapalnej vody zodpovedá za objemový rast buniek a samonosnosť nezdrevnatených častí rastlín (listov, mladých stoniek a pod.). Voda vo vakuole každej z buniek vyvíja zvnútra tlak na bunkovú stenu (turgor). Pri vyrovnaní tlaku vakuoly a odporu bunkovej steny dochádza k stavu tzv. plnej turgescencie bunky. Ak sú bunky turgescentné, vyvíjajú tlak na seba navzájom a pletivá sú pevné. Pri strate vody ochabujú, vädnú. V rade prípadov sú bunkové steny v pletivách usporiadané tak, že v rôznych smeroch viažu molekuly vody rôznymi silami. Zmena obsahu vody potom vedie k pohybom celých orgánov alebo ich častí – napr. zmena polohy listov v priestore a ich zvíjanie, rýchle pohyby listov pri dotyku (Mimosa), otváranie a zatváranie prieduchov, pohyby kvetných častí, otváranie a zatváranie šišiek a tiež zmena polohy celých vetiev v dôsledku prítomnosti tzv. reakčného dreva.
Hodnoty merného skupenského tepla topenia ľadu, varu vody a jej merná tepelná kapacita vysvetľujú mimoriadne účinky vody v termoregulácii rastlinných pletív. Tie časti drevín, ktoré majú vysoký obsah vody (vzrastné vrcholy, dužnaté plody drevín, kambiálna zóna a pod.), majú vysokú teplotnú stabilitu. Spotreba tepla pri výpare vody z listov (transpirácii) je veľmi dôležitá pre ich ochladzovanie. Naopak, pri procese mrznúcia vody sa ochladzovanie pletív spomaľuje. Uvedené objemové zmeny vody počas fáz jej topenia a tuhnutia sú hlavnou príčinou poškodenia rastlinných pletív, najmä pri opakovaných a rýchlych výkyvoch teploty pod a nad 0 °C. Ďalšie schopnosti vody ako reaktantu, donora aj akceptora elektrónov a ľahko pohyblivého rozpúšťadla poukazujú na jej úplne zásadný a nezastupiteľný význam.
Gradienty v dažďových zrážkach, kontrastná expozícia a rozdiely v textúre a hĺbke pôdy, ktoré majú zásadný vplyv na vodnú bilanciu rastlín, majú dopad na vegetáciu, jej typy a funkčný stav. Voda hrá aj podstatnú úlohu pri dekompozícii, zvetrávaní a riadi toky živín v pôde. S vodným režimom je úzko spojená minerálna výživa, fotosyntéza, respirácia, rast aj architektúra rastlinných orgánov.

Vodný potenciál, turgor a osmóza v bunkách stromov
Obsah vody a jej termodynamický stav
Fylogeneticky vzato, život vznikol vo vode a voda zostáva základným médiom, v ktorom prebiehajú biochemické procesy rastlín. Cytoplazma obsahuje v priemere takmer 90 % vody (vzhľadom na čerstvú hmotnosť), na lipidy bohaté organely ako chloroplasty a mitochondrie obsahujú okolo 50 % vody, pokiaľ ide o celé orgány či pletivá rastlín, listy a jemné korene obsahujú okolo 80 %, vodivá časť dreva – biel – okolo 50 %. Nadzemné časti suchozemských (terestrických) rastlín neustále strácajú vodu výparom. V závislosti od ich schopnosti kompenzovať krátkodobé kolísanie obsahu vody a prežívať stratu vody rozlišujeme rastliny na poikilohydrické a homoiohydrické. Poikilohydrické rastliny (huby, niektoré riasy a lišajníky) prispôsobujú svoj obsah vody vlhkosti okolia – majú malé bunky bez centrálnej vakuoly. Pri vyschnutí zmenšia svoj objem a ich životné funkcie sú potlačené, bez toho aby došlo k poškodeniu protoplasty. Homoiohydrické rastliny obsahujú v bunkách vakuóm, ktorý v určitom rozmedzí vyrovnáva vplyv vonkajších vlhkostných podmienok. S vývojom kutikuly, trichómov, riadenia transpirácie prieduchmi, so vznikom suberinizovaných povrchov a prevetrávaných systémov, s vývojom vodivých sústav na dlhé vzdialenosti a s vývojom koreňových systémov zabezpečujúcich dodávku vody, sa stali suchozemské rastliny schopné zachovávať si do určitej miery svoje vnútorné prostredie aj počas náhlych zmien vlhkosti prostredia, a v dôsledku toho sa rozšírili v globálnom meradle. Aj v rámci homoiohydrických rastlín nachádzame fázy, v ktorých sa správajú ako poikilohydrické a znášajú významné straty vody. Sú to zrelé semená, ktoré si udržujú klíčivosť po niekoľko desiatok až storočí tým, že majú schopnosť znížiť životné procesy pri radikálnom obmedzení svojej hydratácie (obsah vody môže byť aj nižší ako 10 %).
Väčšina hydratačnej vody je viazaná v cytoplazme a bunkovej stene, ktorá ju viaže napätím v rozsahu cca 1 až 10 (i viac) MPa, a to v závislosti od hustoty celulóznych fibríl. Sily viažuce vodu ku štrukturálnym elementom matrice (bunková stena, koloidy) možno vyjadriť ako matričný potenciál. Kvantitatívne vzato, význam uloženej vody v bunke stúpa s mierou jej dostupnosti. Najľahšie dostupná voda sa vyskytuje vo vakuolách, t. j. bunkových kompartmentoch špecializovaných ako vodné rezervoáre, alebo v odumretých bunkách slúžiacich ako hydrocyty. Takto je uložená viac ako polovica vody obsiahnutej v listoch. Avšak ani táto voda nie je úplne mobilná, pretože je osmoticky viazaná na rozpustené látky ako cukry, organické kyseliny, sekundárne metabolity rastlín a ióny. Sily, ktoré viažu vodu v roztokoch, sa vyjadrujú ako osmotický potenciál. Iba chemicky čistá voda má najvyššiu špecifickú voľnú energiu, t. j. potenciál vykonávať prácu (napr. rozpúšťať ióny, obklopovať koloidné častice) a voľne sa pohybovať. Vodný potenciál chemicky čistej vody je základom pre vyjadrenie relatívneho chemického potenciálu vody, ktorá je rôznymi silami pútaná v ktoromkoľvek mieste systému pôda – rastlina – atmosféra. U chemicky čistej vody je hodnota jej potenciálu najvyššia, t. j. nulová, u vody viazanej v systémoch sa znižuje, t. j. dosahuje záporných hodnôt. Závisí od vyššie uvedeného osmotického potenciálu (koncentrácie osmoticky aktívnych látok, oligosacharidov v rastline, solí v pôde), tlakového potenciálu (turgorového tlaku na bunkové steny zvnútra buniek a hydrostatického tlaku) a matričného potenciálu (adhézii k bunkovým stenám, vododržnosti koloidov a pod.). Vodný potenciál možno chápať aj ako prácu potrebnú na zvýšenie potenciálu viazanej vody na jeho hodnotu u čistej vody. Zvyčajne sa vyjadruje ako množstvo energie na jednotku objemu (v jednotkách J . m⁻³), jednotku látkového množstva (J . mol⁻¹) alebo ako tlak (v jednotkách MPa).
Osmotický potenciál roztoku klesá nielen s rastúcim počtom rozpustených častíc, ale aj so vzrastajúcou teplotou. Makromolekulárne látky môžu byť prítomné v značných hmotnostných množstvách, bez toho aby osmotický potenciál výrazne znížili. Avšak polymerácia malých molekúl na makromolekuly (napr. premena cukru na škrob a reverzia tohto procesu – hydrolýza) môže osmotický potenciál roztoku silne ovplyvniť, a teda čistý tok vody môže byť rastlinou regulovaný. V protoplastoch s centrálnou vakuolou existuje tesná súvislosť medzi osmoticky viazanou vodou v bunke a dostupnosťou vody v cytoplazme.
Dostupnosť vody je vyjadrená ako celkový vodný potenciál vodného systému (napr. bunky, bunkového kompartmentu alebo vonkajšieho roztoku, zvyčajne označovaný symbolom Ψ), čo znamená, že čím negatívnejší je vodný potenciál daného systému, tým nižšia je dostupnosť vody zo systému pre okolie. Osmotická zložka vodného potenciálu závisí od charakteru a koncentrácie roztoku a je daná Van Hoffovým vzťahom:
kde: R je univerzálna plynová konštanta (8,31 mol⁻¹K⁻¹), T je absolútna teplota (K), ρw je hustota vody, C = Ns/V je celkový počet mólov v roztoku na jednotku hmotnosti vody v symplaste, resp. objemu vody v bunke V, Φ je osmotický koeficient, ktorý zahŕňa neideálne vlastnosti daného roztoku v termodynamickom zmysle.
Celkový vodný potenciál vodného systému zahŕňa k osmotickej zložke vodného potenciálu aj zložku matričného a tlakového potenciálu.



